Život ako Božie dielo
Július Rajčáni
Abstrakt. Genetický kód hrá pre živú bunku rozhodujúcu úlohu. Je to základný biologický princíp, ktorého účelom je uchovať optimálnu usporiadanosť živej hmoty a pritom zabezpečiť jej sústavnú obnovu. Gény skrývajú v sebe program, ktorý zaručuje stabilitu životných procesov a súčasne umožňuje ich zdokonaľovanie v procese evolúcie. Pôvod genetického kódu ako informácie nie je vedecky objasnený. Keďže jeho podstata naráža na hranice absolútna, je súčasťou tajomstva stvorenia. Teológia prírody nás privádza k uznaniu neredukovateľnej zložitosti, ktorá zakladá komplexnú skladbu buniek na základe genetickej informácie prítomnej v chromatíne bunkových jadier, respektíve v chromozómoch deliacich sa buniek. Vo vedeckých kruhoch naďalej pokračuje diskusia, či informačný obsah makromolekúl zakladajúcich život (DNA, RNA ako aj bielkoviny) je výsledkom náhodného usporiadania ich skladbových jednotiek, alebo je následok trvalo pôsobiacej koncepcie. Keď rozumom odhaľujeme skryté zákonitosti života, spoznávame (hoci nie úplne) reálne jestvujúcu harmonickú zložitosť, svedčiacu pre koncepciu aktu stvorenia. Materializmus a agnosticizmus sú ateistické interpretácie, ktoré sa vyznačujú redukciou komplexnej reality. Tento spor odráža protichodné stanoviská v ľudskom myslení, ktoré sa tiahnu dejinami ľudstva. Sekulárna spoločnosť tradujúca protichodnosť vedy a viery sa opiera o prekonaný a zjednodušený výklad, ktorý ustrnul na mechanickom opise životných dejov. Viera v Stvoriteľa je dar ducha, ku ktorému však možno dospieť aj rozumovou úvahou, lebo najnovšie poznatky molekulárnej biológie neodporujú viere v Boha, skôr naopak svedčia v jej prospech.
Abstract. The life as divine agency. The genetic code is essential for each living cell. It is a biological principle determining the optimal structure of cells and allowing long term maintenance of living substance along with its reproduction. The genetic code acts as hidden information governing the processes of life and assuring their inherited stability. Nevertheless, the genetic information undergoes continuous improvement during evolution. Chemical composition of the genetic material as well as the order of its units (nucleotides) within the dsDNA molecule(s) had been widely deciphered. However, the origin of precise succession of nucleotides providing genetic information for proteosynthesis has remained a cardinal philosophical problem, which challenges the limits of our understanding. Natural theology teaches us to accept the non-reducible complexity of the living cell, characterized by the unity of genetic material with the information encrypted in nuclear chromatin and/or chromosomal substance. Even when the human mind is able to understand several aspects of the harmonic complexity of life, novel scientific theories are never fully explanatory, since any discovery opens newly emerging questions. This fact proves the non-exhaustible diversity of creation. Materialism and agnosticism possess the tendency to simplify reality when explaining the succession of nucleotides and the corresponding order of amino acids within cellular proteins by their random combination and selection. The controversy whether genetic code has evolved by chance or whether it is the result of a continuously acting divine program, cannot be solved by solely scientific approaches. The statement that science on one hand and faith in creation on other hand were opposite to each other, results from the attitudes within society. Explanations provided by the atheistic and/or materialistic philosophers are based on old-fashioned mechanistic theories rather than on achievements of contemporary science. The faith in God is still a gift of spirit, but can be widely supported by reason and/or rational thinking, since it does not contradict to but rather rises from the recent discoveries of molecular biology.
Motto: Laici majú svoje ľudské, ako aj profesionálne úsilie prepojiť s náboženskými hodnotami do životaschopnej syntézy. Lebo Boh aj ich povolal k tomu, aby plnili svoje úlohy v duchu evanjelia a aby prispievali k posväteniu sveta zo svojej iniciatívy a vlastným príkladom.
Lumen gentium, II. vatikánsky koncil, 1962 [25]
Úvod
Vedecký výskum živej prírody na molekulovej úrovni dospel v posledných desaťročiach k pozoruhodným poznatkom, ktoré protirečia materializmu, ako ho poznáme zo starších diel biológov a niektorých filozofov na prelome 19. a 20. storočia. Napriek tomuto značná časť vedeckej obce, ako aj vzdelaná sekularizovaná verejnosť je presvedčená, že agnostický postoj k otázkam pôvodu života je moderný. Vo veci evolúcie živých tvorov a pôvodu človeka prevláda názor, že ich vývoj vysvetlený Darwinom poskytuje dosť pádnych argumentov pre materialistické videnie sveta. Dejiny vývoja filozofických postojov od nástupu osvietenstva ukazujú, že materialistický svetonázor a ateizmus nie sú iba neblahým dedičstvom dialektického materializmu násilne presadzovaného v období komunistickej totality ako „vedecký svetonázor“. Ateizmus sa vyskytoval už v antike. V súčasnosti je však pomerne rozšírený najmä v krajinách západnej Európy, hoci tam viaceré generácie vzdelancov žili v podmienkach slobodnej demokratickej spoločnosti. Dôkazom toho je napríklad dielo akademického teológa A. Kee [31], ktorý odporúča dodržovať kresťanskú etiku a morálku aj ľuďom, ktorí v Boha neveria.
Cieľom tohto článku je poukázať na skutočnosť, že súčasná veda a jej objavy neodporujú viere vo vyššiu a všemocnú bytosť, ktorú kresťanské Credo definuje ako „Stvoriteľa vecí viditeľných i neviditeľných“. Ľudia od nepamäti verili v nadprirodzené sily najmä preto, lebo inak si nevedeli vysvetliť pôvod prírody, ktorá ich obklopovala. Súčasné pokroky prírodných vied (najmä biológie a mikrobiológie), ktoré viedli k odhaleniu molekulových základov životných procesov, ukázali, že racionálne uvažovanie o prírode nás privádza späť k uznaniu prvotného významu ideí, plánu alebo rozvrhu, teda nehmotného princípu, moderne označovaného ako informácia [13]. Nadväzujúc na myšlienky sv. Tomáša sa dá predpokladať, že skúmaním usporiadania hmoty je možné dospieť k presvedčeniu o existencii Boha, ktorý je prvotnou príčinou všetkého.
V našej úvahe sa sústredíme na stručný výklad molekulovej podstaty životných procesov v bunkách, v ktorých nepochybne nachádzame organizáciu na základe logického programu. Prírodovedné myslenie a rozumové poznanie sa s rozvojom vedy stali určujúcim základom oblasti teológie, ktorá sa nazýva prirodzená teológia [5, 6, 8]. Prirodzená teológia, niekedy označovaná aj ako „teológia prírody“, považuje existenciu vesmíru, ako aj živých bytostí za dôkaz tvorivej inteligencie ich prvotného pôvodcu, teda Stvoriteľa [37]. Veda o živej prírode (biológia a jej príbuzné disciplíny) podobne ako prirodzená teológia vypovedajú o obdivuhodnom Božom diele. Veda a viera sa doplňujú, lebo predstavujú poznanie tohože bytia odlišnými prístupmi. Veriaci vedec vníma pôsobenie prírodných zákonov ako dôkaz Božej múdrosti a jeho tvorivej dokonalosti. Po II. vatikánskom koncile (1962 – 1965) bol vytvorený priestor pre rekapituláciu poznatkov prirodzenej teológie na dialóg s agnostikmi a so sekularizovanou liberálnou spoločnosťou [25]. Dnes možno viac ako predtým platí, že ...Ozajstná autentická viera neumenšuje slobodu ani rozum, prečo by teda kresťanská viera a rozum sa mali báť jeden druhého [19].
Pohľady na vedu z pozície filozofie
Na začiatku tejto úvahy treba konštatovať, že kresťanstvo umožnilo rozvoj experimentálnych vied v ďaleko väčšom rozsahu než iné monoteistické náboženské kultúry. Opisná veda sa už od 17. storočia usilovala o presný a objektívne platný záznam pozorovanej skutočnosti [26]. Mocným nástrojom výskumu sa okrem merania pri pozorovaniach a presnej dokumentácie nameraných hodnôt stal experiment. Pokus (experiment) sa vyznačuje pozorovaním určitého prírodného javu v presne vymedzených laboratórnych podmienkach, na vhodnom modeli a na základe vopred určeného plánu. Modely majú dvojaký účel: umožňujú vedcom zanedbať v danom kontexte nepodstatné detaily skúmaných objektov začlenených do času a priestoru; zároveň poskytujú zobrazenie aj takých skúmaných objektov, ktoré v danej chvíli nie sú prístupné našim zmyslom [29]. Skúmanie príčiny (kauzality) javu pomocou experimentu sa obvykle sústreďuje na jedinú premenlivú príčinu, a to aj vtedy, ak jestvuje viacero príčin. Pri skúmaní komplexných javov je preto vhodné analyzovať reťazec kauzality z rôznych uhlov pohľadu, iba tak nás vedecké poznanie približuje k pravde [16]. Za kritérium pravdivosti výsledku sa považuje porovnanie modelového pokusu s reálnou skutočnosťou, ktorá existuje v prírode mimo dosahu experimentátora. Získané poznatky majú obvykle tú platnosť, ktorá je definovaná podmienkami príslušného experimentu. Interpretácia výsledku určitého pokusu spočiatku zodpovedá platnej teórii (paradigme). Po období pokojného výskumu, počas ktorého sa hromadia nové publikované fakty, niekedy však nastane náhla (revolučná) zmena hypotézy, ktorá vyvrcholí vo formulácii novej teórie. Akumulácia nových poznatkov umožní formulovať nové teórie hypotézy, lebo nahromadenie doposiaľ nepoznaných faktov vo vede časom vedie k zavrhnutiu starej teórie a ku vzniku novej paradigmy [12]. Nová paradigma poskytuje nádej na prekonanie predchádzajúcich odborných sporov a dáva priestor pre nástup ďalšej fázy výskumu, v ktorom už dominuje nová teória. Vidíme teda, že dejiny poznávania sú výsledkom zdokonaľovania jednotlivých vedeckých poznatkov a prekonávania s tým súvisiacich konfliktov.
Filozofia vedy sa usiluje o syntézu vedeckého poznania na úrovni najvyššie možného stupňa všeobecnosti [17]. Jeho objektom je objasňovanie vedeckých výrokov a ich súvislostí [44]. Keďže veda má veľa špecializovaných odborov, ignorovanie filozofie ako nástroja zovšeobecnenia izoluje súbory vedeckých poznatkov. Pre izoláciu jazyka špecialistov nedochádza k dostatočnej syntéze nových výsledkov ani k ich včleneniu do širších vzájomných súvislostí. Ak výskum iba zakladá navzájom oddelené (a pre nezasvätených vzdelancov málo zrozumiteľné) poznatky, informačná bariéra bráni zaradeniu nových výsledkov bádania do bežného spôsobu myslenia. Prepojenie nových výsledkov vedy s teológiou môže byť zahatané, pokiaľ sa vedci nepokúsia sprostredkovať nové informácie v primeranej forme a na určitej filozofickej úrovni, aby sa mohli stať súčasťou zovšeobecňujúcej teórie alebo hypotézy. Súčasný vedec – špecialista môže byť pragmatik, ktorý nepociťuje potrebu zovšeobecňovať. Pragmatik neuvažuje o pôvode živej hmoty, neskúma ani účel ani spôsob evolúcie života, lebo je presvedčený, že skutočné poznanie sa zakladá iba na pozorovateľných a merateľných faktoch [21]. Už v štádiu skúmania si pragmatik mimovoľne uvedomuje, že nadobudnúť úplné poznanie nie je vlastne ani možné. I keď rešpektujúc túto pravdu [18], vedecký realizmus je prírodovedný a zároveň aj filozofický postoj, ktorý sa usiluje zovšeobecniť poznanie založené na faktoch [30]. Vedec realista je si vedomý toho, že každé ním nadobudnuté poznanie je iba aproximáciou skutočnosti a nepredstavuje definitívny výklad. Ako to už vyplynulo z diskusie o paradigmách, veda je sebakriticky otvorená voči neočakávaným výsledkom.
Každý nový poznatok obvykle vedie k potrebe ďalšieho výskumu. Ak sa výskumník vzdá stratégie neustáleho rozširovania vlastného obzoru a neusiluje sa získať aspoň čiastočné poznatky z príbuzných vedných odborov, ním nadobudnuté výsledky môžu byť aj prameňom nechcených omylov. Príkladom je použitie pojmu atóm na označenie najmenšej stavebnej jednotky hmoty, ktorá sa v chemických reakciách (na rozdiel od molekúl) správa ako nedeliteľná častica. Demokritos vo svojej úvahe o skladbe hmoty vyslovil domnienku, že všetko pozostáva z rovnakých (tých istých) neviditeľných malých skladbových jednotiek, ktoré nazval „atomos“ (nedeliteľný). K definícii nedeliteľnej jednotky hmoty Demokritos dospel rozumovou (v dnešnom poňatí špekulatívnou) úvahou, ktorá mohla byť približne takáto: predstavme si, že delíme nejaký viditeľný objekt, napríklad jablko. Pri veľmi veľkom počte delení dospejeme k jednotke, ktorá sa už ďalej deliť nedá. Častica, ku ktorej pri takomto pomyslenom delení dospejeme, však už nebude jablko, ale „atomos“, z čoho sa skladá aj čokoľvek ostatné. Pri správnej konfrontácii Demokritovej úvahy s dnešnými poznatkami vedy pojmu „atomos“ najlepšie zodpovedá častica kvark. Z kvarkov, ktorých existencia bola overená experimentálne, sa ako z nedeliteľných jednotiek skladajú protóny a neutróny. Tie tvoria jadrá atómov všetkých prvkov zaradených do Mendelejevovej sústavy [23, 40].
Filozofia vedy, ale najmä prirodzená teológia, napomáhajú sprostredkovať optimálnu interakciu vedy s teológiou Zjavenia. Vieme, že interakcie vedeckého a teologického poznania môžu byť (a v dejinách aj boli) rozličné [39]. Konflikt je nežiaduca forma, ktorej príkladom by mohol byť prípad Galilei. Sväté ofícium napriek Galileovmu pozorovaniu pohybu mesiacov Jupitera okolo tejto planéty považovalo popularizáciu Kopernikovho heliocentrického modelu pohybu planét za unáhlené a nevhodné [42]. Na tomto mieste sa žiada pripomenúť, že myšlienka o jedinečnosti planéty Zem napríklad z pohľadu vzniku života neprotirečí vedeckej argumentácii. Predstava o jedinečnosti Modrej planéty v našej Galaxii, kde boli (Božím plánom) vytvorené podmienky na vznik života, vôbec neznamená, že v rámci slnečnej sústavy neplatí princíp kopernikovského usporiadania. Nezávislosť sa zakladá na tom, že hoci obe, veda aj teológia, interpretujú prírodu, pôsobia ako celkom oddelené disciplíny, pričom sa každá z nich rozvíja vlastnou cestou. Ignorovanie možného nesúladu však neprispieva k riešeniu problémov vynárajúcich sa pri navzájom sa prekrývajúcich témach. V porovnaní s dvoma uvedenými formami vzájomnej interakcie je sľubnejšia asimilácia, t. j. hľadanie spoločných pojmov, ktoré umožnia budúci dialóg. Na tento postup nadväzuje vlastný dialóg, dotýkajúci sa styčných oblastí oboch disciplín, ktoré by bolo možné postupne riešiť za podmienky vzájomného rešpektovania odlišných východísk a pri hľadaní adekvátnych prístupov. Pravda, ktorú vedci hľadajú, je vlastne atribút Boha, nás ľudí teda nekonečne presahuje a nik z nás si pravdu nemôže privlastniť. Môžeme sa k nej blížiť v tej miere, v akej v sebe čo najautentickejšie rozvíjame podobu Boha, na ktorú sme boli stvorení. To je dôležitá myšlienka, ktorá rezonuje v encyklike Benedikta XVI., že totiž k pravde sa môžeme približovať po rozličných cestách ako viery, tak aj rozumu. Encyklika zdôrazňuje, že tieto cesty sú nielenže vzájomne kompatibilné, ale komplementárne: vyžadujú jedna druhú! „Rozum sa neprestajne potrebuje očisťovať vierou. No aj náboženstvo sa potrebuje stále očisťovať rozumom“ (1, čl. 56, Caritas in veritate). V tejto situácii vzťah medzi vierou a vedou nemôže byť zápasom o „územia“ či „sféry vplyvu“, akýmsi bojom o moc, ale neustálym hľadaním vzájomného dialógu v prospech vlastného napredovania [32]. Medzi otázky spoločného záujmu teológie a biologických vied by podľa názoru autora tejto eseje mohla patriť aj jednota materiálnej a informačnej povahy genetickej substancie. V ďalšom texte autor rozvíja myšlienku, že genetická informácia, ako základ organizácie živých tvorov, pochádza od Boha. Prírodovedná argumentácia autora sa pokúša uchopiť tému postupujúc zdola nahor (zo žabej perspektívy), kým teológia pristupuje k analýze prírodných javov zhora nadol (vtáčia perspektíva). Z úspešného dialógu vyplynie konsonancia – nachádzanie súladu v oblastiach spoločného záujmu za predpokladu plného rešpektovania ďalšieho autonómneho rozvoja oboch disciplín.
Genetický kód a jeho význam
Genetika je náuka o dedičnosti. Jedným zo zakladateľov tejto vednej disciplíny bol Johan Gregor Mendel (1822 – 1884), kňaz a opát augustiniánskeho kláštora v Brne. Mendel na základe presne vymedzených dedičných znakov rastlín hrachu, ktoré štatisticky sledoval cez dve generácie (F1 a F2), formuloval základné zákony dedičnosti (potvrdili ich o 30 až 50 rokov neskôr Morgan, Johannsen a Bates). Uvedení vedci formulovali pojem génu, čo je jednotka prenášajúca genetické znaky. Gén je materiál (chemicky definovateľná látka), ale aj informácia zároveň. Jeho informačný obsah sa vzťahuje na zloženie určitej bielkoviny, ktorú kóduje. Genetický kód (kodón) signalizuje informáciu pre každú jednotlivú aminokyselinu, ktorá je skladbovou jednotkou bielkovín, vrátane jej poradia v reťazci bielkoviny. Ak bielkovina definovaná génom má povahu enzýmu katalyzujúceho určitú kľúčovú chemickú reakciu, tak platí vzťah jeden gén – jeden enzým. Ak je gén pre príslušný enzým porušený (a nedôjde k jeho kompenzácii pomocou génu druhého pohlavného partnera), tak u dieťaťa môže vzniknúť defekt v štruktúre príslušného enzýmu. To sa prejaví ako „vrodená“ choroba, ktorého príčinou je nefunkčný gén.
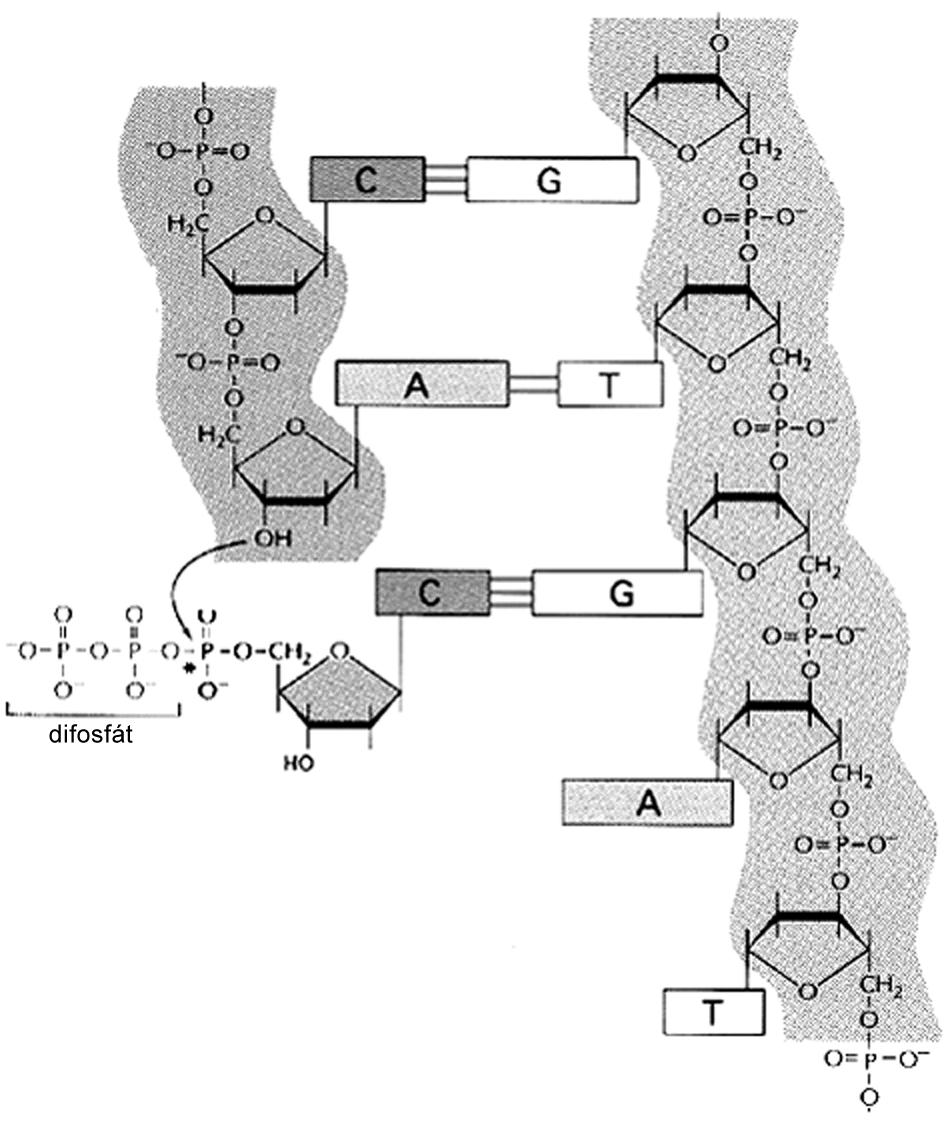
Watson a Crick [43] objasnili skladbu dvojitej špirály dvojvláknovej DNA (dsDNA = deoxyribonukleová kyselina, ds = double strand). Vyslovili predpoklad, že gény sú lokalizované na jednom z vláken dvojitej špirály. Ako vidno na obr. 1, každé vlákno dsDNA sa skladá zo 4 druhov chemických jednotiek(nukleotidov), ktoré sa označujú skratkou A, T, G a C (podľa mena príslušných báz). Navzájom sú komplementárne vlákna dvojitej špirály DNA prepojené pomocou vodíkových mostíkov, pričom sa naproti sebe ležiace nukleotidy navzájom párujú podľa ustáleného princípu (A:T alebo G:C). Pri syntéze nového vlákna DNA činnosťou zložitého enzýmu zvaného DNA polymeráza sa jednotlivé nukleotidy prikladajú vždy tak, aby naproti ležiace bázy tvorili párované dvojice. Tým sa pri každom delení tvorí také isté nové vlákno, ktoré je akoby „odliatkom“ na matrici starého vlákna [podľa 1]. Celá molekula dsDNA u človeka pozostáva približne z 3.2x109 nukleotidov (skladbových jednotiek), ktorých poradie je presne určené. Pomocou chromozómov, do ktorých sa vlákno dsDNA pre delení bunky organizuje, sa molekula dsDNA, a tým aj genetická informácia prenesú na potomstvo. Z veľkého počtu nukleotidov molekuly dsDNA tvoria gény najviac 5 %. Gény predstavujú vysoko konzervatívne úseky DNA; v molekulárnej genetike (respektíve v molekulárnej biológii) sú označované ako čítacie rámce (ORF, open reading frame). Svojbytnosť jednotiek ORF v molekule dsDNA potvrdilo aj sekvenovaníe celej ľudskej DNA. Zistilo sa tiež, že úseky v sekvencii dsDNA (obvykle priradené tesne pred jednotlivými ORF) tvoria nekódujúce regulačné sekvencie (vysvetlené ďalej). Vlastnú genetickú informáciu v úseku ORF tvoria tzv. kodóny (obr. 2), ktoré pozostávajú z trojitej kombinácie nukleotidov. Každý kodón je akoby signálnym znakom morzeovky, lebo určuje niektorú aminokyselinu v reťazci polypeptidu (bielkoviny). Systém kodón/antikodón (na kódujúcom vlákne DNA, na molekule mRNA a tRNA, porov. obr. 3) umožní zachovať informáciu o poradí aminokyselín pri syntéze bielkovín. Poradie aminokyselín v bielkovine napokon zodpovedá poradiu kodónov „prepisovaného“ čítacieho rámca na vlákne DNA.
Kódujúce vlákno DNA predstavuje molekulu s vysokým informačným obsahom („inteligentná molekula“), v ktorom je poradie jeho zložiek (nukleotidov) presne stanovené [39]. Jednotlivé ORF vo vlákne DNA predstavujú základné štruktúry pre prenos genetickej informácie na príslušnú bielkovinu na strane jednej, ako aj na potomstvo (chromozómy pre delení buniek) na strane druhej. Pri prepise genetickej informácie (génu) obsiahnutej v čítacom rámci (ORF) sa z molekuly DNA vytvorí komplementárna molekula mRNA (enzým RNA polymeráza). Molekula mRNA nesie tú istú informáciu, hoci chemicky predstavuje iný druh molekuly (obr. 3). Molekula mRNA sa od DNA líši jednak cukrom (ribóza, nie deoxyribóza), ale aj jednou odlišnou bázou (U = uridín namiesto T = tymín). Pri prepise molekuly mRNA sa nukleotid T prítomný v DNA páruje s nukleotidom A v molekule mRNA, ale A v DNA sa páruje s U v mRNA.
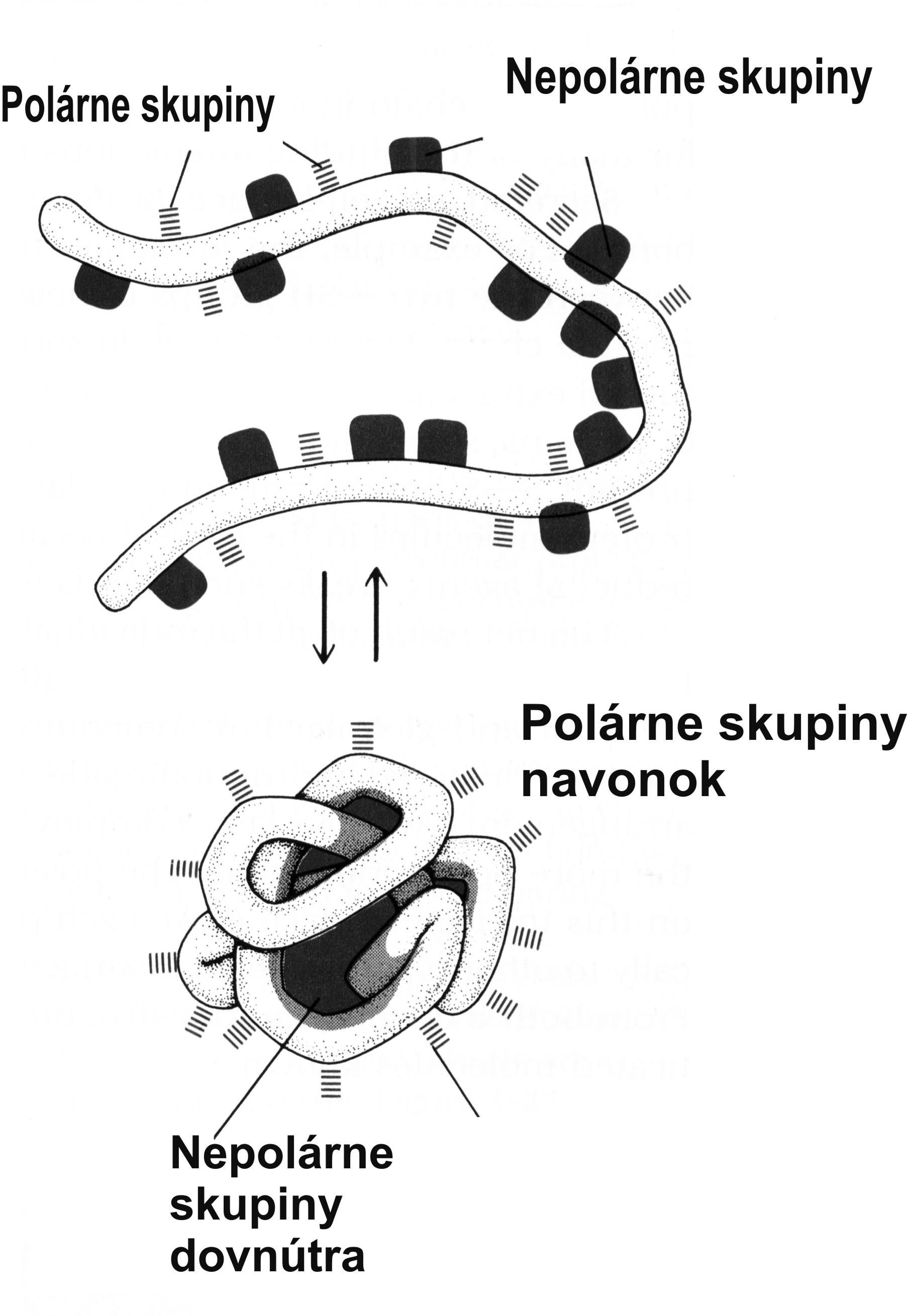
Makromolekula bielkoviny je zložená z jednotiek zvaných aminokyseliny. Aminokyseliny majú kyslú karboxy skupinu (–COOH) na jednom, a zásaditú amino skupinu (–NH2) na druhom konci. Mnohé aminokyseliny majú viac ako tieto dve polárne skupiny, takže zvyšná nezreagovaná kyslá alebo zásaditá skupina vo vode disociujú (polárne skupiny). Jednotlivé aminokyseliny sú spojené väzbou CONH (peptidová väzba). Reťaz väzieb CONH tvorí pevné lano s bočnými výčnelkami odlišnej polarity, resp. s nepolárnymi hydrofóbnymi skupinami (obr. 4A). Na základe elektrostatických síl, resp. schopnosti príslušnej oblasti (peptidu) zmiešať sa s vodou dlhé „lano“ polypeptidu sa vo vodnom prostredí stočí do klbka (obr. 4B) alebo vytvorí iný trojrozmerný útvar (mreža, špirála, vrkoč a pod.). Bielkoviny sa skladajú najmenej zo 100, ale najviac z 1 500 aminokyselín, ktoré v rozličných kombináciách tvoria molekuly odlišných vlastností. Ako som už uviedol, každá aminokyselina má v molekule bielkoviny svoju presnú polohu, určenú poradím príslušných kodónov v čítacom rámci kódujúceho vlákna DNA. Podobne ako DNA, aj bielkovina je „inteligentná“ molekula s vysokým informačným obsahom. Pri všetkých troch uvedených makromolekulách (DNA, RNA, polypeptid/bielkovina) základom ich informačného obsahu je zmysluplné poradie skladbových jednotiek, čo určuje ich funkčný význam.
Funkčne významné a často nezastupiteľné poradie nukleotidov v DNA, ako aj z toho plynúce poradie aminokyselín v bielkovinách je dané informačným obsahom, ktorý sa podľa ateistickej koncepcie vytvoril náhodne [22, 35]. V ďalšej časti poukážeme na nízku pravdepodobnosť náhody ako určujúceho faktora vzniku práve tých bielkovín, ktoré sa ustálili v priebehu evolúcie živých foriem [27]. Prirodzená teológia predpokladá, že usporiadanie aminokyselín v bielkovinách je súčasťou Božieho plánu stvorenia. Hnacou silou evolúcie bielkovín nebola náhoda, ale koncepcia (respektíve program) obsiahnutá v Božom pláne. Presvedčenie o riadiacom programe síce vylučuje, že by základným riadiacim činiteľom evolúcie mohla byť náhoda, ale nevylučuje, že by náhoda pôsobila ako súbežný vonkajší faktor modifikujúci evolučný proces (vysvetlené ďalej).
Teologický význam genetického kódu je v rozpoznaní jednoty hmoty a informácie. Informácia o poradí aminokyselín v bielkovinách je zabudovaná do matérie DNA. Táto informácia je objektívnou realitou, ktorú dialektický materializmus podobne ako aj viacerí filozofi 19. a 20. storočia celkom prehliadli. Základom dedičnosti je genetický kód, ktorý je obsiahnutý vo vláknach chromatínu (pri bunkovom delení v chromozómoch), pozostávajúcich z dlhej molekuly dsDNA zvinutej zvláštnym spôsobom v bunkovom jadre. Dedičnosť pôsobí ako endogénny faktor, ktorý podmieňuje našu reakciu na vonkajšie prostredie, na individuálne životné podmienky každého jedinca. V súhre so životným prostredím dedičná danosť determinuje naše zdravie a telesné vlastnosti, ktoré môžu byť vystavené náhodnému pôsobeniu rozličných vonkajších podmienok.
Úvahy o bunke ako organizovanej živej hmote
Náuka o živote (všeobecná biológia) zastrešuje rad špecializovaných odborov výskumu (molekulová biológia, mikrobiológia, virológia, imunológia, fyziológia, patológia, genetika, ale aj anatómia, histológia a pod.). Bunky rastlín aj živočíchov (na rozdiel od buniek baktérií) majú oddelené jadro, ktoré obsahuje chromatín (špeciálne zvinuté vlákna dsDNA). Jadrá buniek sú centrály pre syntézu a prepis RNA (transkripcia opísaní zhora). Bunka „vyrába“ bielkoviny pre vlastné potreby, alebo ich pre potreby organizmu vylučuje navonok. Syntéza proteínov nastáva na ribozómoch. Ribozóm je miesto, kde sa tvoria reťazce bielkovín na základe prekladu genetického kódu obsiahnutého v molekule mRNA (zhora opísaná translácia, obr. 3). Bunky obsahujú bielkoviny tvoriace ich kostru, ale i ďalšie bielkoviny, ako sú enzýmy, ktoré katalyzujú chemické reakcie. Enzým urýchli chemickú reakciu, ktorá je náročná na spotrebu energie i na časovú kinetiku. V neprítomnosti enzýmu by mnohé reakcie, ktoré sa v bunkách odohrávajú, nemohli vôbec uskutočniť. Princípom funkcie enzýmu je schopnosť aktivovať molekuly substrátu, ktoré do reakcie vstupujú pri syntéze zložitejších molekúl. Komponenty budúcej zlúčeniny sa k sebe tesne priblížia, aby nadobudli aktivačnú energiu (obr. 5); umožní to konfigurácia záchytného miesta pre substrát. Pri zlučovacej reakcii sa napokon uvoľní viac energie, než bolo použité na aktiváciu reakčných zložiek. Iné enzýmy, naopak, zložité látky rozkladajú, teda vykonajú opačnú reakciu, akou bola ich syntéza. Lysozym je príkladom enzýmu, ktorý rozkladá disacharidy, resp. polysacharidy v prítomnosti vody (hydrolýza), čím vznikajú jednoduché cukry (monosacharidy). Lysozym funguje na princípe priestorovej konfiguračnej zmeny. Hydrolýza nastane len vtedy, ak sa reťazec polysacharidu ohne do určitého uhla. Konformačnú zmenu substrátu sprostredkuje enzým pomocou svojho záchytného miesta. Špecifické reakčné miesto enzýmu potom umožní vlastný rozklad substrátu (di- alebo polysacharidu) pridaním molekuly vody.
Poznáme stovky ba tisíce enzýmov, ktoré sú základom chemických pochodov, súhrnne označovaných ako bunkový metabolizmus. Enzýmy sú súčasťou zoskupení, ktoré umožnia komplexné reakčné pochody. Polyfunkčné enzýmové systémy predstavujú neredukovateľnú zložitosť (unreducible complexity). Príkladom sú navzájom prepojené oxidačné a fosforylačné pochody v bunkách cicavcov, ktoré sa uskutočňujú na organelách zvaných mitochondrie. Molekuly viacerých cytochrómov, ktoré tvoria „dýchací reťazec“ aj u aeróbnych baktérií, prenášajú elektrón, čím dochádza k postupnému využitiu energie uvoľnenej v spojitosti s redukciou kyslíka. Prvým „odberateľom“ elektrónu z atómu kyslíka je enzým cytochrómoxidáza. Je to veľmi stará molekula, ktorú aeróbne baktérie využívajú vyše jednej miliardy rokov. Na mitochondrách cicavčích buniek je pridružená ATP syntáza, ktorá pracuje ako protónová (H+) pumpa. ATP sa neustále tvorí z ADP, čím sa „skladuje“ energia do makroergnej fosfátovej väzby molekuly ATP. Štiepenie ATP je zdrojom energie pre rozličné enzýmové reakcie, napríklad aj pri práci svalov. Naše svaly získavajú energiu z rozkladu cukrov v prítomnosti ATP. Cicavce preto spotrebujú veľa kyslíka, pričom vylučujú CO2 a H2O. Rastliny vybavené chlorofylom spotrebujú CO2 a za pomoci energie slnečného žiarenia spätne tvoria organické látky (ako napríklad sacharidy).
Bunka je základná jednotka života, schopná samostatnej existencie. Každá bunka sa tvorí iba z inej bunky. Jej prvotný vznik z neživej organickej hmoty je hlbokým tajomstvom, ktoré veda doposiaľ neodhalila. Predpokladá sa, že aby sa chemické reakcie, potrebné na kopírovanie prvotnej RNA pomocou primitívneho enzýmu (RNA polymeráza) mohli vôbec udiať, museli sa obe makromolekuly nachádzať vo vnútri priestoru obaleného bunkovou membránou (obr. 6). Molekuly enzýmu RNA polymeráza vstupovali do prvotných interakcií s molekulami RNA až vtedy, ak sa mohli k sebe dostatočne priblížiť. Bez membrány ohraničujúcej priestor budúcej bunky by pravdepodobnosť bezprostredného kontaktu oboch molekúl v prvotnom mori bola bývala minimálna. Bunková membrána sa postupne vyvinula v orgán, ktorý je viac ako len bariéra vymedzujúca priestor pre životné reakcie. Membrána asociovala s viacerými špeciálnymi transportnými bielkovinami, ktoré pumpujú ióny a organické látky smerom dovnútra ako aj von z bunky (obr. 7). Stabilné chemické zloženie cytoplazmy, ktorú umožňuje bunková membrána, sa nazýva homeostáza. Homeostáza je základným predpokladom životných dejov katalyzovaných enzýmami, ktoré môžu prebiehať iba v stabilných podmienkach vnútorného prostredia.
Pri životných procesoch uvedených v tabuľke 1 sa jednoduchá následnosť (jedna príčina a jediný dôsledok) uplatňuje len v niektorých prípadoch. Jednoduchá následnosť môže pribúdaním nových enzýmov vytvoriť uzavretý okruh, v ktorom reakcie neustále pokračujú. V takomto prípade nemusí byť spočiatku jasné ani to, kde je začiatok (prvotná príčina) a kde je koniec (posledný dôsledok). Pribúdaním ďalších enzýmov a ich pomocných bielkovín vznikajú reťazce následností, v ktorých jeden dôsledok môže mať niekoľko príčin (aby jav nastal, musia byť splnené všetky podmienky), alebo jediná príčina môže vyvolať viacero dôsledkov. Následnosť javov v bunke nie je vždy priamočiara aj preto, že tam pôsobí zložitá sieť interakcií s viacerými možnosťami vzájomných vplyvov. Pri prerušení jednej z reťazí kauzality alternatívne reťazenie prevezme náhradnú funkciu (časté v mozgu pri interakciách medzi nervovými bunkami, ale pri imunitnej obrane alebo metabolických poruchách). Interpretáciu príčiny a následku v živej bunke teda znejasňujú komplikované interakcie medzi molekulami bielkovín a nukleových kyselín, ktoré môžu byť natoľko zložité, že sa sústavne objavujú ich nepoznané detaily. Pri odhaľovaní zatiaľ ešte nepoznaných súvislostí musíme filozofovať opatrne. Nemožno zamieňať nepoznané s naozaj nepoznateľným tajomstvom, ktoré je za končenými hranicami absolútneho poznania.
Vieme, že hmota (matéria) je zložená z elementárnych častíc, ktorými sú kvarky (skladbové jednotky protónov a neutrónov) a elektróny. Z nich pozostávajú atómy prvkov, ako aj väčšie molekuly. Hmota podlieha fyzikálnym zákonom, medzi jeho základné vlastnosti patria zotrvačnosť, hmotnosť a existencia v priestore i v čase. Hmota mení svoje formy; premieňa sa na energiu, ale jej celková masa nezaniká. Hmota je nositeľkou chemických reakcií a elektrických javov. Základnou vlastnosťou hmoty definovanej ako matéria je jej objektívna existencia mimo nášho vedomia. Nezávisle od nášho vedomia však v skutočnosti jestvuje aj živá substancia, ktorá má svoju usporiadanosť a informačný obsah. Usporiadanosť aminokyselín ako stavebných jednotiek bielkovín sa zakladá na informácii zapísanej v genetickom kóde. Kopírovaním dcérskej DNA na molekule materskej DNA sa informácia uchováva pre ďalšie generácie. Na presnosti poradia nukleotidov (podľa kodónov) záleží najmä v úsekoch čítacích rámcov jednotlivých génov. Pri reprodukcii buniek v dcérskej DNA sa predsa len môžu objaviť malé odchýlky (bodové mutácie). Rozsiahlejšie zlomy alebo straty sekvencií (delécie) sa tiež vyskytujú pri niektorých chorobách, táto problematika však nie je predmetom našej úvahy. Bodové mutácie buď navodia poruchu, alebo niekedy umožnia skvalitniť funkciu bielkoviny. Mutácie, ktoré prispôsobujú funkciu na nové podmienky, napomôžu smerovaniu príslušného organizmu k dokonalejším formám. Antropický princíp [29] predpokladá, že parametre hmotných častíc určujúce vlastnosti atómov vodíka, uhlíka, kyslíka a dusíka (a ďalších prvkov živej hmoty) mohli byť od samotného počiatku vzniku vesmíru nastavené tak, aby mohli neskôr byť využité pri vzniku života (slabý antropický princíp). Informácia o výstavbe „inteligentných“ makromolekúl (zložených z organických zlúčenín na báze uhlíka, kyslíka, dusíka, síry a fosforu) mohla byť od samotného počiatku uložená v substancii ako vopred určený program (silný antropický princíp).
Ak by život vznikol a vyvíjal sa iba neriadeným (náhodným) spôsobom, teda pri odmietnutí koncepcie Bohom riadeného programu stvorenia, vystúpia do popredia viaceré, pre súčasnú vedu neriešiteľné problémy (uvedené v tabuľke 2). Dajú sa vyjadriť dvomi základnými okruhmi otázok: problém vzniku prvej aminokyseliny (je ich 20 druhov) a časová náročnosť vzniku funkčných makromolekúl, ktoré by vzišli z náhodného usporiadania aminokyselín do „správneho“ poradia. Teoreticky pri každom proteíne by mohla reagovať každá aminokyselina s ktoroukoľvek zo zvyšných 19 aminokyselín, čo by sa opakovalo toľkokrát, koľko má tá ktorá bielkovina aminokyselín. Napokon ostáva ešte jeden ďalší nemenej významný „háčik“: spôsob organizácie živej hmoty do bunkovej štruktúry. Vedci sa zaoberali aj otázkou, ktorá kódujúca molekula jestvovala skôr: DNA, či RNA? Pravdepodobný sled dejov sa v dávnej minulosti nepodobal tomu, čo sa deje v súčasnosti. Dnes je už málo častý spôsob kopírovania informácie smerom od RNA do DNA (reverzná transkripcia RNA do DNA), hoci sa predpokladá, že práve tento spôsob mohol byť prvotný. Uvedený archaický sled kopírovania genetickej informácie „využívajú“ niektoré vírusy (retrovírusy), ktoré pritom spôsobujú v bunkách nemalé škody, často vedúce aj k nádorovej premene. Genómy, ktoré vo víruse majú sekvenciu RNA, sa v bunke pomocou opačne kopírujúceho enzýmu (zvaného reverzná transkriptáza) prepíšu do DNA sekvencie (tzv. provirus). Provírus sa včlení do DNA hostiteľskej bunky ako nový úsek. Mnohé retrovírusy sa zachovali už len ako nekompletné fragmenty dávno predtým integrovaných provírusov. Preto sa predpokladá, že spätne prepísané sekvencie DNA hrali určitú, v súčasnosti nie celkom objasnenú úlohu v evolúcii živých organizmov (pozri ďalej).
Tabuľka 2 Záhady biológie, ktoré hraničia s absolútnym poznaním
|
1. Ako vznikli prvé aminokyseliny? Pokus o syntézu jednoduchej aminokyseliny: predchodca glycínu z CH2O a HCN [metán + čpavok + voda a vodík, v prítomnosti silného elektrického výboja, Miller, 1953, citované podľa 3] doposiaľ nebolo úspešné.* 2. Usporiadanie aminokyselín do „správneho“ poradia pri tvorbe prvých bielkovín. Pri jednoduchšom polypeptide je pravdepodobnosť správneho usporiadania aminokyselín 10161 : 1 [Guye, CE, 1919, citované podľa 3]. Pri použití všetkých 20 aminokyselín, ktoré sa môžu umiestniť v ľubovoľnom poradí v bielkovine s 500 jednotkami, sa teoreticky predpokladá až 20499 variácií. Pri tak vysokom počte možností je pravdepodobnosť užitočného výsledku náhodného usporiadania mimoriadne nízka. K tomu pristupuje aj časová náročnosť týchto procesov pre potrebu „testovania“ funkčnosti každej zo vzniknutých molekúl. 3. Pôvod tripletov (kodónov), ktoré svojím poradím v čítacích rámcoch jednotlivých génov (v molekule DNA, resp. RNA) určujú poradie aminokyselín v polypeptidoch. Poradie nukleotidov v génoch (ORF), ako aj kodóny signalizujúce aminokyseliny sa podľa ateistov ustálili spontánne, na základe náhodných variácií (trial and error, pokus a omyl). Problémy uvedené v bodoch 1, 2 a 3 sa ešte znásobia pri ich vzájomnom prepojení do interakcií vedúcej k zmysluplnej funkčnosti. 4. Pôvod tzv. regulačných (promotor) sekvencií DNA, ktoré sú dôležité pre prepis (expresiu) génov V tejto úvahe nie je priestor na podrobnú diskusiu o regulácii prepisu mRNA, čiže spôsob expresie jednotlivých génov. Jestvujú sústavne prepisované gény, gény prepisované na povel (vonkajší signál) a gény, ktoré sú v diferencovanej bunke s určitou funkciou natrvalo utlmené. 5. Princípy stavby enzýmov a ich usporiadanie do funkčných systémov. 6. Vznik prvotných buniek: tvorba mikropostredia ohraničeného lipidovou membránou. 7. Princíp diferenciácie buniek mnohobunkových organizmov (od zárodku k novorodeniatku: všetky bunky tohože jedinca majú tie isté gény, ale rozličné bunky obsahujú rozličné bielkoviny. Treba podotknúť, že diferencované bunky tohože organizmu exprimujú približne osminu alebo desatinu všetkých proteínov, ktoré by mohli prepísané z ich utlmených génov (porov. bod 4).** |
* Opakované pokusy zostrojiť aspoň čo i len tú najjednoduchšiu aminokyselinu (glycín) neboli zatiaľ úspešné.
** Niektoré bielkoviny sa syntetizujú len za určitých podmienok, napr. po indukcii stimulačnými látkami; takto bunky odpovedajú na vonkajšie podnety (signály).
V primitívnych bunkách sa čoskoro objavili bielkoviny, ktoré sa viažu na DNA, a spolu s kľúčovým enzýmom pre kopírovanie mRNA (RNA polymeráza) umožnili proces regulovaného prepisu (expresie) génov. Už bolo uvedené inde, že všetky bunky tohože jedinca obsahujú rovnakú DNA. Takže v každej bunke tohože jedinca sú prítomné všetky gény, ktoré tvoria jeho genofond. Selektívny prepis niektorých bielkovín v bunkách so špeciálnymi funkciami umožnilo ich diferenciáciu a je aj príčinou ich nerovnakého vzhľadu. Spoločný predchodca rôznorodých buniek organizmu však môže byť prítomný aj v dospelosti ako nediferencovaná pluripotentná (kmeňová) bunka, schopná diferencovať sa rozličným smerom. Len nedávno boli identifikované tzv. kmeňové bunky dospelých, schopné dodatočnej diferenciácie, ktoré v prípade potreby nahradia náhle poškodené tkanivo. Celá problematika tzv. kmeňových buniek, ako aj otázky okolo odlišnej expresie génov v rozlične diferencovaných bunkách mnohobunkového organizmu presahuje rámec tohto článku.
Napriek rozsiahlemu pokroku vedeckého poznania každý výskum naráža na problém hraníc poznateľnosti najmä z pohľadu absolútna, ktoré prináleží iba Všemohúcemu. Teológia (vrátane teológie prírody) predpokladá, že absolútne poznanie je vlastné iba Bohu. Naša snaha hlbšie poznávať realitu životných dejov a prírody vedie iba k tomu, že sa pred výskumníkmi vynárajú vždy nové, doposiaľ neznáme súvislosti, ktoré je potrebné ďalej analyzovať. Tým dochádza k silnej diverzifikácii vedných odborov, čo sa napokon stáva prekážkou k pochopeniu obecných princípov skúmaného systému. Teológia prírody odmieta taký výklad reality, ktorý vyžaduje redukciu kauzality, alebo vytvára trvalé zjednodušujúci výklad neredukovateľnej komplexnosti biologických dejov. Neredukovateľná zložitosť predstavuje systém pozostávajúci z mnohých navzájom dobre prispôsobených súčastí, v ktorom však principiálna zmena spôsobí narušenie funkcie celku [4]. Výklad, ktorý smeruje k zjednodušeniu neredukovateľnej zložitosti na nižšiu úroveň reálnej skutočnosti je síce možný z dôvodu pedagogickej zrozumiteľnosti, ale ako koncepcia oberá životné deje o ich informačný rozmer. Metóda redukcie komplexnej zložitosti sa nesmie stať podkladom trvalej teórie či hypotézy. Už aj scholastici dávneho stredoveku postrehli, že princípom života nie je telo samotné, ale spôsob jeho fungovania (sv.Tomáš Akvinský). Každé bytie (teda aj živá hmota) nadobúda príčinu svojej existencie mimo seba, teda nie v sebe samom [24]. Pojem stvorenia (creatio) naznačuje, že forma hmoty a usporiadanosť substancie by neexistovali mimo Boha, ale že vznikli skrze neho. Už sv. Tomáš píše, že svet osôb, zvierat a rastlín je ako kniha (moderne „informačná knižnica DNA“), do ktorej Stvoriteľ zapísal pravdu o tvoroch samých a taktiež v nej zanechal stopy o sebe. Pri materialistickom výklade sa živá bunka redukuje na konglomerát chemických reakcií; ľudské myslenie sa mení na komunikáciu medzi nervovými bunkami pomocou hmotných signálov (transmiterov). Fyziologický materializmus redukuje životné pochody na chemické interakcie a fyzikálny pohyb [10, 14].
Tabuľka 2 vymenúva ťažko riešiteľné výskumné ciele, ktoré v súčasnosti predstavujú hranice biologického poznávania. Autor sa vopred sa ospravedlňuje všetkým, ktorí predpokladajú, že problémy v nej uvedené bude možné vedecky vyriešiť bez toho, že by bolo potrebné prekročiť hranice absolútneho poznania. Pokus o výpočet zásadných medzier vo vedeckom poznaní, ktoré by mohli slúžiť ako argument na podporu prirodzenej teológie, vôbec neznamená, že každý nevyriešený biologický problém predstavuje nepoznateľnú skutočnosť. Nie je totiž dôvodné predpokladať, že každý nepoznaný jav zakladá nepoznateľné tajomstvo Stvoriteľa. Kritici „teórie medzier“ právom poukazujú na to, že pokrokmi vo vedeckom bádaní sa neustále redukuje rozsah neprebádaných oblastí reality. Mnohé javy, ktoré pri starej paradigme (pôvodne platnej) nebolo možné vysvetliť, sa stávajú zrozumiteľnými po nastolení novej paradigmy. Žiadne ľudské poznanie však nemôže prekročiť hranice absolútneho poznania. Preto vedci, ktorí sa zaoberajú zásadnými otázkami okolo vzniku bunky a pôvodu života, neustále narážajú na hranice poznania, cez ktoré sa nevedia (a pravdepodobne ani nemôžu) dostať. Naše poznávanie sa iba približuje k pochopeniu podstaty javov, avšak dokonalé vzájomné pochopenie všetkých súvislostí ostáva Božím tajomstvom. Treba preto odlíšiť témy, ktoré veda zásadne nevie (ba ani nemôže) vyriešiť, od takých problémov, ktoré z rozličných príčin veda doteraz ešte nevyriešila [20]. Teóriu medzier (gaps) by bolo preto vhodnejšie považovať za teóriu hraníc poznávania. Teória hraníc poznania by mohla byť dobrým námetom na spoluprácu teológie a filozofie vedy. Sem patrí definovanie limitov neredukovateľnej zložitosti, ktorými sa veda približuje k zásadne nepoznateľnému absolútnu. Pripomínam tu citát z úvodného prejavu Jána Pavla II. na vedeckej konferencii vo Vatikáne z roku 1988: „Veda sa snaží zbaviť náboženskú vieru nánosov povery; viera však môže zbaviť vedu nenáležitej absolutizácie a falošného modloslužobníctva hmoty“ [34].
Skúmanie substancie ako objektívnej reality vyžaduje racionálne myslenie, ktoré odhaľuje jej vnútorné zákonitosti. Tie vyjadrujú výsledky matematiky, fyziky, chémie a biológie. Poznateľnosť reality takého vysokého stupňa aktivitami mozgu vyžaduje integrálne spojenie medzi rýdzo fyzikálnym (elektrické a chemické signály), ako aj duchovným (nehmotným) princípom. Keďže určité oblasti mozgu môžu byť sídlom myslenia (ako duševnej činnosti), je zrejmé, že ich základom nemôže byť len rýdzo fyzikálna a chemická realita (hmota), ktorá vôbec nie je vybavená informačným obsahom. Mozog človeka nie je iba autonómny počítač [3]. Ak obrazom sveta je usporiadanosť, ktorú spoznávame pomocou rozumu, nemôžeme súčasne tvrdiť, že myslenie je iba náhodným produktom neriadeného „pohybu“ hmoty, ktorá počas svojej nekonečnej existencie nepodliehala nijakému vyššiemu organizačnému princípu (mindless matter). Racionálne myslenie nemôže byť založené iba na pohybe chemických molekúl a ich interakciách s bunkovými receptormi, ktoré samy osebe nie sú zárukou nijakej racionality. Aby sa nejaký úsudok (assertion) mohol považovať správny alebo nesprávny, musí mať racionálny pôvod. Avšak nijaká fyzikálna hmota (ani tá, z ktorej sa skladá mozog), by sama osebe nemohla byť zdrojom uvažovania, pokiaľ nepredpokladáme, že jestvuje taký informačný (duchovný) obsah, na ktorom sa zakladajú životné procesy vrátane tých, na ktorých spočíva ľudské myslenie [33].
Evolúcia a prirodzená teológia
Prekvapujúci súhlas vedeckých výsledkov a dialektického myslenia je pre cirkev významným argumentom, lebo evolúcia je už viac ako hypotéza [citované z prejavu Jána Pavla II. na Valnom zhromaždení Pápežskej akadémie vied, 1996, 15]. Darwin (1809 – 1882) kedysi napísal, že malé zmeny v génoch živých bytostí (v dnešnej nomenklatúre bodové mutácie) vedú k novým formám [7], ktoré sú lepšie prispôsobené na podmienky vonkajšieho prostredia (prirodzený výber). Pri každom delení bunky sa nová molekula DNA tvorí iba na matrici (podľa vzoru) starej molekuly, čo je základný princíp dedičnosti. Chyby párovania nukleotidov v čítacích rámcoch sa vždy neprejavia závažnou zmenou aminokyseliny (mutácia). Niektoré bodové mutácie (malé zmeny) môžu znamenať selekčnú výhodu, pretože navodia kvalitnejšiu funkciu (vznikne nový genetický variant, prípadne aj nový druh). Pri reprodukcii živých tvorov stoja proti sebe dva opačne pôsobiace mechanizmy. Na jednej strane môže nastať výhodná selekcia molekuly zmenenej bodovou mutáciou génu, na druhej strane pri bunkovom delení sa uskutoční kontrola génu, ktorá „zisťuje“, či sa pri syntéze novej DNA vytvorila absolútne presná kópia pôvodnej sekvencie. Mikroorganizmy, ako aj bunky vyšších organizmov sú vybavené mechanizmom, ktorý udržuje relatívnu stabilitu sekvencie konzervatívneho genetického kódu kontrolou prekopírovaných sekvencií čítacích rámcov (génov). Kontrolu správnosti zabezpečuje sústava enzýmov, ktorá pri každom delení identifikuje poradie nanovo pridaných nukleotidov v čítacích rámcoch. Po ukončení fázy S (syntéza DNA) bunkového delenia ale tesne pred presunom chromozómov do dcérskych buniek (fáza G2) sa v konzervatívnych sekvenciách čítacích rámcov pomocou sady reparačných enzýmov eliminujú prípadné chyby (nesprávne nukleotidy, mismatch). Po splynutí zárodočných buniek nového potomstva sa rovnaké rodičovské chromozómy k sebe prikladajú na dobu viacerých hodín (konjugácia), aby sa presunmi alebo výmenami celých úsekov DNA (aj viacerých génov) mohli vzájomne „opraviť“ chybné miesta. Keby tieto mechanizmy kontroly génov nejestvovali, ľudia by húfne umierali na nádory a rozličné vrodené chyby. Mechanizmus nápravy chýb pri kopírovaní génov znižuje pravdepodobnosť mutácií sto až tisícnásobne, čo tiež možno pričítať k Božiemu plánu na udržanie zdravého potomstva. Opísaný mechanizmus na celé milióny rokov stabilizoval dôležité makromolekuly, najmä také kľúčové enzýmy, ako je zhora uvedená cytochrómoxidáza.
Mutácie, ktoré nastali mimo konzervatívnych úsekov DNA (mimo génov), sa nemôžu prejaviť zmenou kodónu, lebo mimo čítacích rámcov kodóny nie sú. Niektorí bádatelia predpokladajú, že mutáciami zmenené čítacie rámce ostávajú naďalej prítomné v DNA ako tzv. redundantné (zdvojené) a zdanlivo nadbytočné nefunkčné gény. Niekedy sa zdvojené gény presunú na iné miesto pomocou pohyblivých elementov, tzv. transpozónov. Takýmito elementmi sú úseky dávnejšie včlenených provírusových sekvencií (uvedené zhora). Redundantné sekvencie by mohli byť svedectvom „vývoja“ genofondu, hoci ich skutočné funkcie zatiaľ nie sú podrobnejšie známe. Fenomén reduplikácie (zdvojenia) génu sa uplatní aj pri tvorbe nových alternatívnych génov (pomocou rekombinačných procesov), čím vznikajú sekvencie kódujúce nové príbuzné bielkoviny. Dnes je zrejmé, že popri malých zmenách (bodových mutáciách) opísaných Darwinom boli pre evolúciu určujúce rozsiahle presuny sekvencií, prípadne výmeny celých dlhších reťazcov DNA. V molekulovej genetike sa tieto deje označujú ako rekombinácia, konjugácia, komplementácia, transpozícia, resp. translokácia. Najviac sú takéto zmeny DNA prebádané u baktérií, pretože baktérie sa v štádiu exponenciálneho množenia rozdelenia skoro každú hodinu. Za života jednej až dvoch ľudských generácií sme sa stali svedkami princípu prirodzeného výberu najmä pri baktériách, čoho príkladom je ich prispôsobenie sa antibiotikám (rezistentné kmene). Odolné kmene väčšinou vznikli presunmi celých génov alebo aj ich skupín (prenos tzv. plazmidmi kódujúcimi enzýmy, ktoré rozkladajú antibiotiká). Podobne aj nové vírusy (vtáčia alebo prasacia chrípka, SARS) vznikli presunmi génov alebo ich preskupením (reassortment). Vidíme teda, že bodové mutácie môžu byť síce významné pre premenlivosť druhov, nepredstavujú však hlavný mechanizmus evolučných zmien, ktoré viedli k novým rodinám alebo čeľadiam organizmov.
Na základe deistickej koncepcie (deizmus) by život mohol byť dôsledkom jednorazového aktu stvorenia [36]. Týmto aktom bola vytvorená hmota, ktorá mohla byť vopred predurčená pre vznik života (antropický princíp). Uvedený výklad možno prijať s tou výhradou, že život je skôr výsledkom trvalého pôsobenia Stvoriteľa (creatio continua). Všimnime si, že aj kniha Genesis hovorí o postupnom stvorení. Ak jednotlivé „dni“ v texte knihy Genesis nahradíme časovo neutrálnejším slovom „etapa“, tak pred našimi očami sa rozvinie poetický opis vývoja našej planéty od stvorenia zeme, jeho oceánov a súše, cez objavenie sa živých tvorov až po človeka. Tento opis v zásadných črtách obstojí aj pred vedeckou analýzou pomocou modernej geológie, paleontológie či antropológie. Tu povedal Boh: buď svetlo a bolo svetlo (Gen 1, 3). Svetlo sa pravdepodobne objavilo celkom na počiatku, teda asi 200 miliónov rokov po veľkom tresku (big bang) pri vzniku prvých primitívnych hviezd (asi pred 13,5 miliardami rokov). Potom Boh povedal: Vody... zhromaždite sa na jedno miesto a ukáž sa súš! A stalo sa tak (Gen 1, 9). Kontinentálna platňa zeme je hrubá 50 – 100 km a je stará asi 3,8 miliárd rokov. Všetky kontinenty spočiatku tvorili jeden celok zvaný Pangea, ktorý oblieval prvotný oceán. Tu Boh povedal: vody, hemžite sa množstvom živých tvorov a okrídlené tvory, lietajte ponad zem. A Boh stvoril veľké morské zvieratá a všetky živočíchy, ktoré sa hemžia vo vode... (Gen 1, 20). Prvé známky života vo forme primitívnej bunky (napr. sinice) sa objavili v prvotnom oceáne pred vyše 3 miliardami rokov. Na začiatku prvohôr (pred 600 miliónmi rokov) sa datujú prvé mnohobunkové organizmy (napr. trilobity). Na začiatku druhohôr, teda asi pred 200 miliónmi rokov, žili v moriach veľké ryby, najmä žralokovité a dvojdyšné ryby. Na súši sa objavili veľjaštery, ktoré opúšťali vody a vo vzduchu lietali prvé vtáky. Biblia jasne uvádza, že život vznikol v prvotnom mori a aj ona pripomína starší pôvod vtákov. Búrlivé premeny kontinentov, erupcie vulkánov a iné katastrofy pred približne 60 miliónmi rokov prežili iba plazy, niektoré druhy bezstavovcov a primitívne cicavce (len asi 10 % vtedajšieho genofondu). Vtedy, na začiatku treťohôr, začína prudký rozvoj cicavcov. Na konci ich vývoja sa napokon objavil najstarší predchodca človeka (asi pred 1 – 2 miliónmi rokov, podľa niektorých vývoj hominoidov trval až 5 miliónov rokov). V knihe Genesis k tomu možno čítať toto: Boh urobil živú zver podľa svojho druhu, dobytok a plazy podľa svojho druhu... (Gen 1, 25). A stvoril Boh človeka na svoj obraz, ...ako muža a ženu ich stvoril (Gen 1, 27). Existenciu dnešného človeka (homo sapiens sapiens, na ktorého sa biblický citát najpravdepodobnejšie vzťahuje, môžeme datovať nanajvýš na desaťtisíc rokov).
Z knihy Genesis nevyplýva nijaký vedecký poznatok. Jej text napriek tomu jasne hovorí, že život sa vyvíjal a zdokonaľoval, a že živé tvorstvo (obr. 8) nebolo spočiatku totožné s tým dnešným. Vznik života je teda suverénny stvoriteľský akt Boha, ktorý všetko, čo stvoril, považoval za dobré [34]. A Boh videl, že je to dobré... (Genesis). Z knihy Genesis priamo nevyplýva, že vývoj života trval niekoľko miliárd rokov, ani že je výsledkom zložitých fyzikálnych, chemických a biologických dejov, ktoré vytvorili podmienky pre jeho vznik [11]. Súčasní kresťanskí evolucionisti vychádzajú z presvedčenia, že evolúcia prebiehala tak, ako ju veda postupne objavuje. Nedávajú stvorenie a evolúciu do protikladu, ale chápu evolúciu ako postupnú, nepretržitú stvoriteľskú činnosť Boha [42]. Evolúcia môže znamenať, že svet je ešte stále neukončený, alebo je prinajmenej v pohybe, lebo sa neustále zdokonaľuje. Evolúcia, ako ju spoznáva veda, je dobrou správou pre teológiu, keďže pripúšťa možnosť neustálych premien smerujúcich k väčšej dokonalosti. Deisti naproti tomu predpokladali, že Boh svet stvoril, potom už doňho viac nezasahoval [36].
Adaptáciu organizmov pomocou prirodzeného výberu (Darwinov princíp), možno považovať iba za súčasť širšieho tvorivého programu, ktorý permanentne ovplyvňuje evolúciu. Pri evolúcii života sa striedali zásadné skoky s pomalým zdokonaľovaním tvorstva. Tento poznatok predpokladá viaceré opakované zásahy tvorivej sily Stvoriteľa (semideizmus), ale nevylučuje ani sústavnú Božiu starostlivosť. Zmeny v génovom vybavení (hromadenie nefunkčných génov, ich presuny a opätovné využitie) v náznakoch vysvetľujú spôsob vzniku principiálne odlišných organizmov (nové rodiny, kmene a čeľade). Konštrukcia úplne novej, odlišnej formy (napríklad vznik cicavcov) predstavuje takúto radikálnu zmenu [9]. Podstatný skok v procese evolúcie sa dopĺňal prirodzeným výberom, keďže genetický kód neposkytoval iný, ako už vopred vymedzený priestor pre tvorbu druhov a ich prispôsobovanie sa podmienkam [2]. Prirodzený výber fixoval náhodné priaznivé zmeny, najmä pokiaľ boli v daných podmienkach výhodnejšie pre ďalšie prežitie (princíp adaptácie). Výhodnejšie formy sa stali súčasťou trvalého „zápisu“ v konzervatívnom genofonde. Takýmto spôsobom nastala drobná diferenciácia v rámci rodov a druhov, čo najlepšie vysvetľuje ich variabilitu. Prirodzený výber by však sám osebe nebol mohol usmerniť evolúciu všetkými možnými smermi, keďže vývoj založený len na náhodných mutáciách by bol neurčitý ako celok.
Čokoľvek Boh mienil programom evolúcie a nech sú výsledky prirodzenej selekcie akékoľvek, kritika teizmu založená na sledovaní života v prirodzených a nedotknutých ekosystémoch (napr. na Galapágoch) hovorí, že sa Boh v nich javí indiferentný, pripúšťa plytvanie a pri ustálení potravinového reťazca neprejavuje nijakú starostlivosť o svoje tvorstvo [28]. V skutočnosti Boh v prírode neplytvá, len zabezpečuje prežívanie druhov pomocou „nadsadenej“ hojnosti. Jeho starostlivosť sa prejavuje najmä pri veľkorysej reprodukcii, ktorá podlieha dynamickým zákonitostiam selekcie. Pôsobenie náhodného výberu na úrovni jednotlivých biologických foriem možno pripustiť aj bez odmietnutia existencie nadprirodzenej bytosti (Boha), pokiaľ nie je spochybnená zásada existencie ním stanoveného programu. Boh pripúšťa prirodzený výber, lebo funguje ako spätná väzba, ktorá na základe prispôsobivosti na existujúce podmienky (adaptácia) zdokonaľuje živé formy. Prirodzený výber iba vyberá optimálnu formu z jestvujúcej „ponuky“ , vyplývajúcej z geneticky daných možností. Dobrým príkladom je napríklad tvorba protilátok na základe endogénne vytvorenej variability v oblasti molekúl imunoglobulínov, ktorá viaže antigén. Konfigurácia antigén viažucej oblasti protilátok je výsledkom génových kombinácií a je daná dedične. Ak sa vnútorne zdedená ponuka stretne s reálnou potrebou určenou vonkajšími vplyvmi (infekcia), vtedy sa začnú množiť práve také klony B-buniek, ktoré pripravujú protilátky príslušnej špecifickosti (klonová selekcia). Uvedený príklad dobre vysvetľuje aj pôvod účelnosti, ktorú nachádzame v živej prírode.
Literatúra
[1] Alberts, B. et al.: Molecular biology of the cell, 5th ed., Garland Science, New York, 2008, 1268 s., 2012.
[2] Asher, R. J.: Evolution and belief, confessions of a religious paleontologist, Cambridge University Press, 2012.
[3] citát Blackmore, C. The mind machine, 1988, In: The neurosciences and the soul, materiál z prednášky prof. Carolla W. F. z Oxfordskej univerzity, Bratislava, 28. 11. 2005.
[4] citát Behe, M. Darwin´s black box: biochemical challenge in evolution, 1996, In: Creation and evolution, materiál z prednášky prof. Carolla W. F. z Oxfordskej univerzity, Bratislava, 24. 10. 2005.
[5] citát Boedder, B. Natural Theology, 1891, In McGrath, A. E.: Přirozená teologie, s. 420 – 426, Blackwellova encyklopédie moderního křesťanského myšlení, Návrat domu, Praha, 2001.
[6] citát Cleobury, F. H. Return to natural theology, 1967, In McGrath, A. E.: In: Přirozená teologie, s. 420 – 426, Blackwellova encyklopedie moderního křesťanského myšlení, Návrat domu, Praha, 2001.
[7] citát Darwin, C. h. The origin of species, 1859, In: McGrath, A. E.: Biologie a křesťanské myšlení, s. 41 – 46, In: Blackwellova encyklopédie moderního křesťanského myšlení, Návrat domu, Praha, 2001.
[8] citát Derham, W. The Physico-Theology, 1713, In McGrath, A. E.: Přirozená teologie, s. 420 – 426, In: Blackwellova encyklopédie moderního křesťanského myšlení, Návrat domu, Praha, 2001.
[9] cituje Haught, J. Deeper than Darwin: The prospect for religion in the age of evolution, 2003, In: Evolutionary biology and the challenges to Philosophy and Theology, materiál z prednášky prof. W. F. Carolla z Oxfordskej univerzity, Bratislava, 10. 10. 2005.
[10] citát Holbach, P. H. D. Systém prírody, In Furstová, M., Trinks, J.: Svet ako organizovaná hmota, s.157 – 164, In: Filozofia, SPN, Bratislava, 1996.
[11] citát Kaufman, G. J. Am Acad Religion, jún 2001, In: Evolutionary biology and the challenges to Philosophy and Theology, materiál z prednášky prof. W. F. Carolla z Oxfordskej univerzity, Bratislava, 10. 10. 2005.
[12] cituje Kuhn, T. S. Štruktúra vedeckých revolúcií, 1957, In Furstová, M., Trinks, J.: Evolúcia vedenia s. 111 – 135, Filozofia, SPN, Bratislava, 1996.
[13] citát Mason, F. The great Design (1932), In McGrath, A. E.: Biologie a křesťanské myšlení, s. 41 – 46, In: Blackwellova encyklopédie moderního křesťanského myšlení, Návrat domu, Praha, 2001.
[14] citát de LaMettrie, J. O. In Furstová, M., Trinks, J.: Svet ako organizovaná hmota, s. 157 – 164, In: Filozofia, SPN, Bratislava, 1996.
[15] cituje Osservatore Romano 24. 10. 1996, In Lobkowitz, M. Evoluční teorie a víra ve stvoření, Teologický týdenník 1, 17 – 19, 2006.
[16] citát Popper, K. R. Logika výskumu,1934, In Furstová, M., Trinks, J.: Evolúcia vedenia s. 111 – 135, Filozofia, SPN, Bratislava, 1996
[17] citát Spencer, H. Systém syntetickej filozofie, 1896, In Furstová, M., Trinks, J.: Evolúcia vedenia s. 111 – 135, Filozofia, SPN, Bratislava, 1996.
[18] citát podľa sv. Pavla z Tarzu „Lebo teraz poznávame len sčasti a len sčasti prorokujeme. Ale keď príde to, čo je dokonalé, prestane, čo je len čiastočné“, List Kor. 13, 9 – 10.
[19] BENEDIKT XVI., cituje Tomko J. „Rozšíriť rozum“: veda a viera, Prednáška laureáta pri udelení ceny Fides et Ratio, 13. 9. 2012, Radosť a nádej, č. 1 – 2, s. 54 – 57, 2012.
[20] cituje Tracey, T. Particular providence and the God of gaps, s. 289 – 324, In: Chaos and complexity, 1995, materiál z prednášky, Divine agency and the autonomy of nature, prof. Caroll W. F. z Oxfordskej univerzity, Bratislava, 14. 11. 2005.
[21] Comte, A.: Kurz pozitívnej filozofie, s. 15 – 18, In: Malá antológia z diel filozofov, II, SPN, Bratislava, 1998.
[22] Crick, F.: Of molecules and man, University of Washington Press, Seattle, 1966.
[23] Glenville, A. R.: Scientifica, Sprievodca svetom dnešnej vedy, Fortuna Libri, Bratislava 2009.
[24] Fotta, P.: Stvorenie sveta podľa Tomáša (filozofická interpretácia stvorenia z ničoho). Radosť a nádej č. 1, 2003, s. 110 – 117.
[25] Foresi, P.: Laici a teológia (úvaha o jednej z veľmi aktuálnych tém cirkvi), Viera a život VI, č. 3, s. 255 – 262, 1996.
[26] Furstová, M. Trinks, J.: Evolúcia vedenia, s. 111 – 135, In: Filozofia, SPN, Bratislava, 1996.
[27] Hoyle, F.: The inteligent universe, London, Michael Joseph ltd, 1983.
[28] Karaba, M.: Cesty k poznaniu Boha, písomný materiál k prednáške UTV v Bratislave, dňa 22. 2. 2006.
[29] Karaba, M.: Základné aspekty používania modelov v prírodných vedách a v teológii. Radosť a nádej č. 2, 2008, s. 31 – 40.
[30] Karaba, M.: Vedecký realizmus ako východisko pre riešenie problému Božej činnosti vo svete. Radosť a nádej č. 1, 2010, s. 15 – 25.
[31] Kee, A.: The way of transcedence; christian faith without belief in God. A Pelican Book, Penguin books, Harmondsworth, Middlesex, UK, 1971.
[32] Košč, S.: Dialóg medzi teológiou, filozofiou a prírodnými vedami vo svetle encykliky Caritas in veritate, Radosť a nádej, č. 1, s. 10 – 14, 2010.
[33] Lewis, C. S.: Is Theology Poetry? 1988, zdroj Wikipedia, Argument from Reason http://en.wikipedia.org/wiki/argument_from_reason
[34] Lobkowitz, M.: Evoluční teorie a víra ve stvoření, Teologický týdenník 1, 17 – 19, 2006.
[35] Monod, J.: „Chance and necessity“, Random House, Vintage Books, New York, 1972.
[36] McGrath, A. E.: Deizmus, s. 72 – 74, In: Blackwellova encyklopédie moderního křesťanského myšlení, Návrat domu, Praha, 2001.
[37] McGrath, A. E.: Přirozená teologie, s. 420 – 426, In: Blackwellova encyklopédie moderního křesťanského myšlení, Návrat domu, Praha, 2001.
[38] Pearcy, N. Thaxton, CB.: Duše vědy: Proměny ve vztahu vědy a náboženství, Návrat Domu, Praha, 1997, 304 s.
[39] Polkinghorne, J.: Věda a teologie: úvod do problematiky, CDK (Centrum pro demokracii a kulturu), Brno, 2002, 170 s.
[40] Rees, M.: Vesmír: unikátna obrazová encyklopédia, Ikar, Bratislava, 2006.
[41] Spišiaková, M.: Spor Galilea Galileiho s Katolíckou cirkvou ako začiatok nového vzťahu medzi profánnymi vedami a teológiou. Radosť a nádej č. 1, s. 20 – 45, 2011.
[42] Tiňo, J.: Stvorenie a evolúcia, Radosť a nádej, č. 2, s. 62 – 66, 2003.
[43] Watson, J. D., Crick, F. H. C.: Molecular structure of nucleic acids. A structure for deoxyribose acid. Nature 171, 737 – 738.
[44] Wittgenstein, L.: Filozofické skúmania, s. 47 – 49, In: Kiczko L. a kol.: Malá antológia z diel filozofov, SPN, Bratislava, 1998.
Obr. 1 Princíp párovania ako základ tzv. komplementarity navzájom zvinutých reťazcov dvojitého vlákna DNA. Uvedený princíp a z neho plynúcej komplementarita (forma a odliatok) je základným princípom tvorby (syntézy) nového vlákna DNA. Nové vlákno DNA nevzniká náhodným prikladaním nukleozidov. Jednotky A, T, G a C sa prikladajú tak, že princípom párovania sa vždy kopíruje ich poradie na už jestvujúcom vlákne. Poznámka: nukleozid sa prikladá prvou fosfátovou skupinou na hydroxyl (–OH) cukru. Po odštiepení dvoch fosfátov sa do vlákna DNA začleňuje príslušný nukleotid.
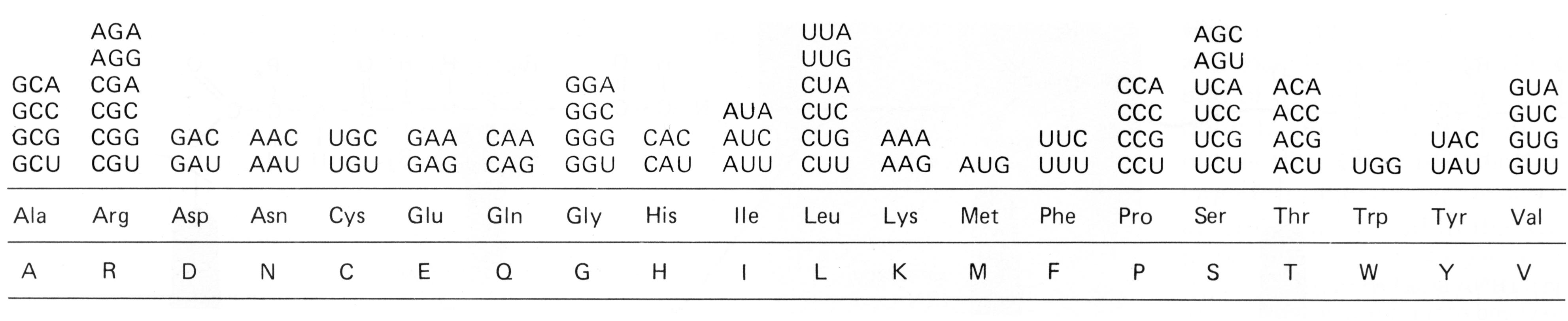
Obr: 2 Kodóny a im zodpovedajúce aminokyseliny. Kodónov je 64, aminokyselín je 20, takže väčšine aminokyselín zodpovedajú najmenej dva kodóny. Začiatočná aminokyselina je M (metionín), ktorému zodpovedá jeden tzv. iniciačný kodón. Tri „stop“ kodóny na schéme uvedené nie sú.
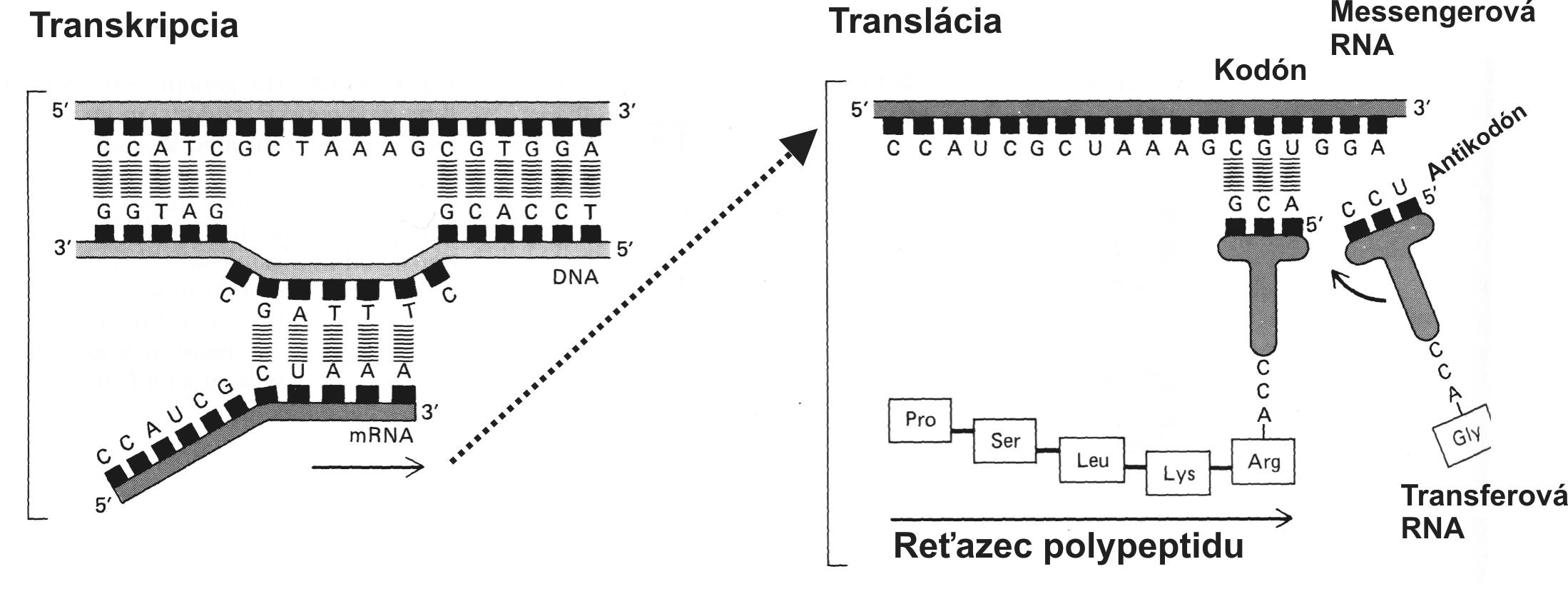
Obr. 3 Princíp prepisu génu (genetickej informácie) na molekulu poslíčkovej RNA (m = messenger RNA). Prepis (transkripcia, transcription) sa odohráva v jadre cicavčej bunky, kde sa nachádzajú zvinuté vlákna dsDNA (chromatín). V mieste rozvoľnenia dvojitej špirály DNA sa prepisuje mRNA a potom sa presúva do cytoplazmy, kde je prítomný aparát na tvorbu bielkovín (ribozómy). Tu sa pomocou iných druhov molekúl RNA (tRNA, t = transfer) tvoria vlákna polypeptidu (bielkoviny). Aminokyseliny sa prikladajú v poradí stanovenom genetickou informáciou obsiahnutou v kódujúcom géne. Táto fáza prenosu genetickej informácie sa nazýva preklad (translácia). Jazykom (kryptogramom) prekladu genetickej informácie obsiahnutej v géne (DNA) na informáciu v molekule bielkoviny sú kodóny, akási morzeovka v „jazyku“ génov (porov. obr. 2). Podľa nej molekuly tRNA prikladajú jednotlivé aminokyseliny. Identifikácia kodónu pracuje na princípe párovania; rozpoznávajúca oblasť na molekule tRNA sa nazýva antikodón.
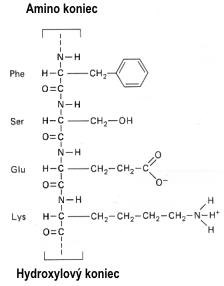
Obr.
4 Princíp tvorby peptidu (obr. 4A), v ktorom sú molekuly
aminokyselín spojené peptidovou väzbou (C=ON–H). Peptidy tvoria
dlhé reťazce (polypeptidy). Pri interakcii s vodou sa polypeptid
stočí v dôsledku vlastností jeho aminokyselín (polárne alebo
nepolárne skupiny). Celá konfigurácia molekuly bielkoviny (obr. 4B),
ktorú vo vodnom prostredí zaujme, je v podstate určená
genetickou nformáciou (kódom). Osvedčené molekuly existujú vyše
miliardy rokov a sú svedectvom dlhého vývoja života a vzájomnej
príbuznosti živých tvorov (porov. obr. 5).
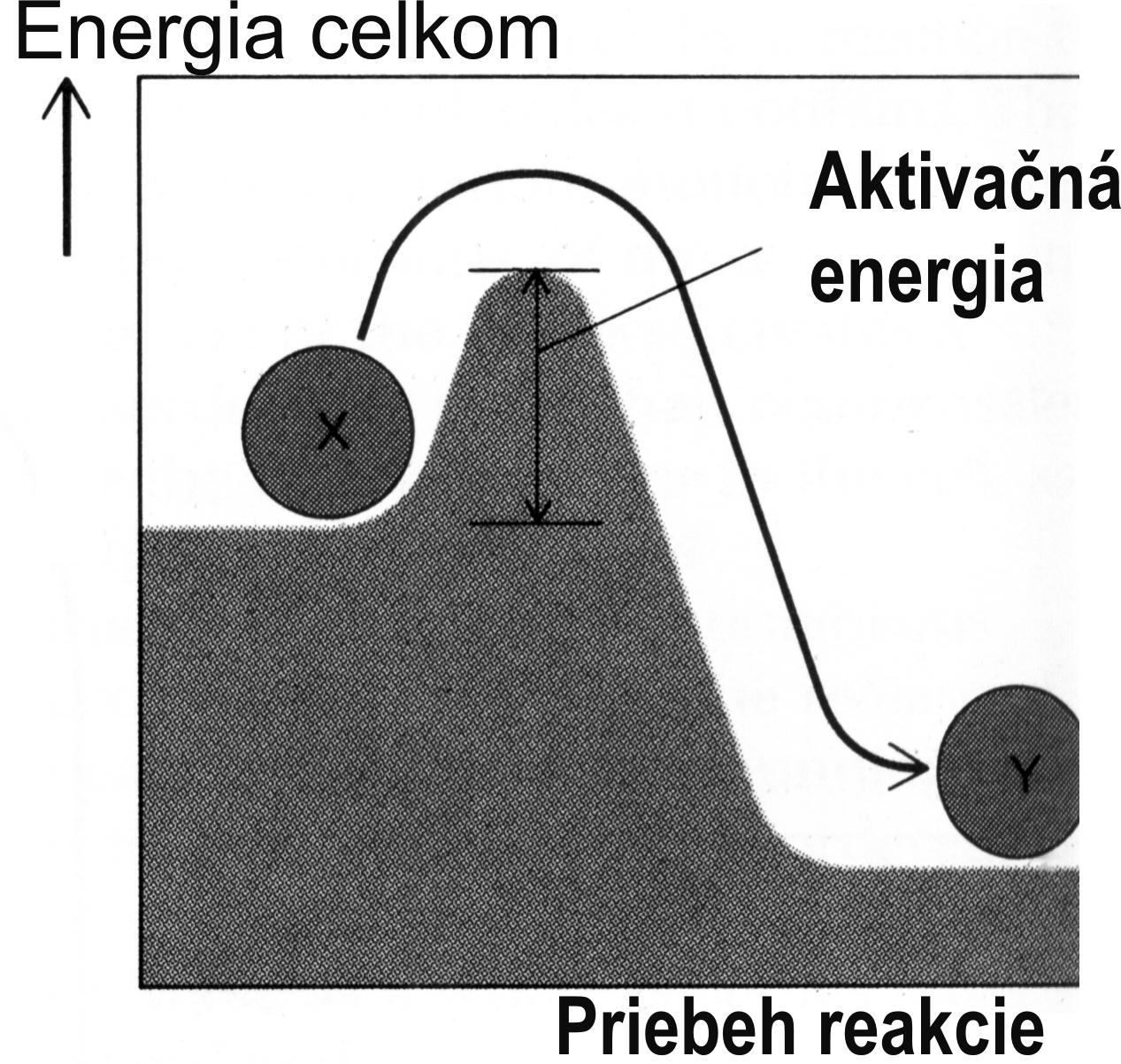
Obr. 5 Enzým ako katalyzátor chemickej reakcie vyžadujúcej aktivačnú energiu.
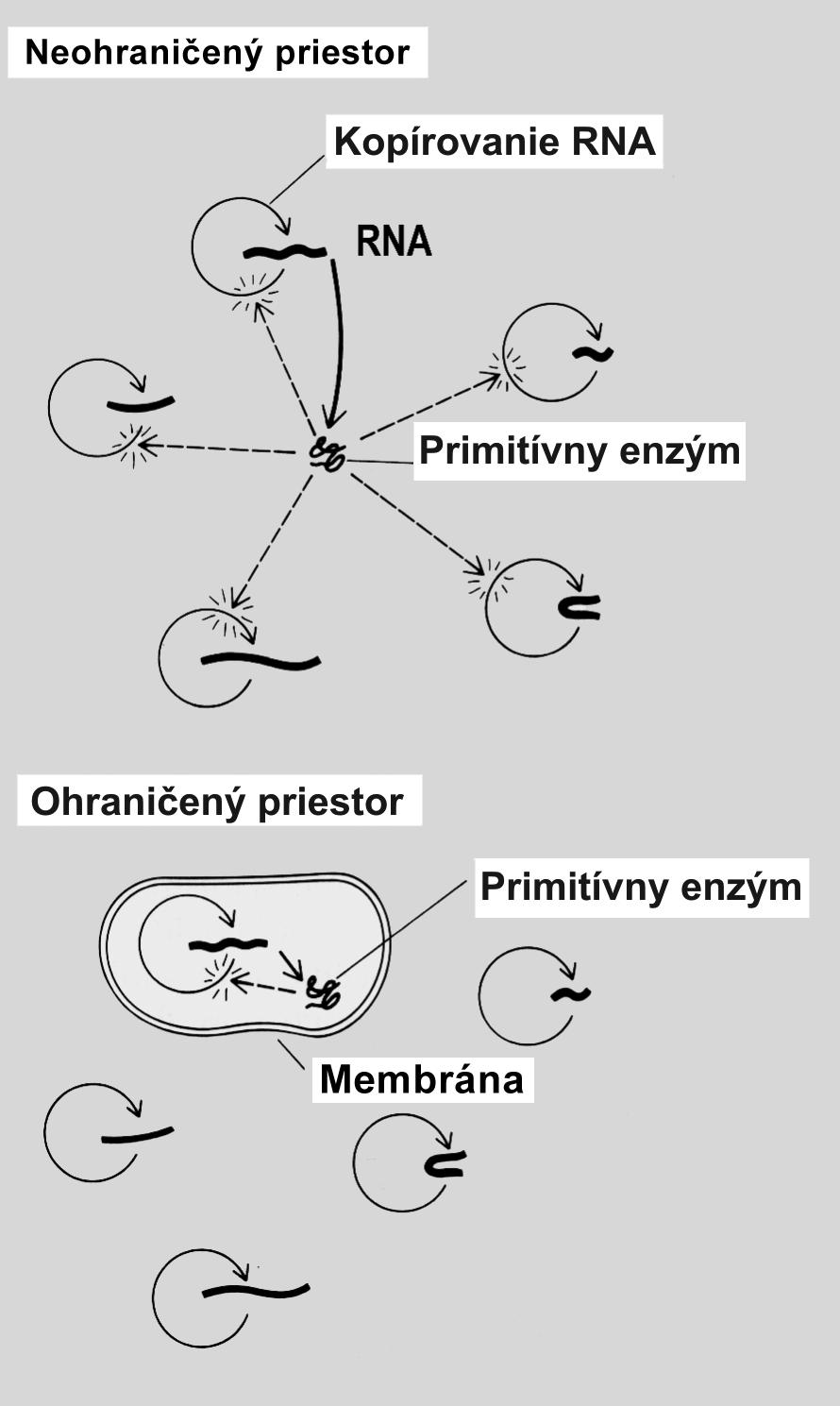
Obr. 6 Schéma interakcie genetickej informácie (obsiahnutej v RNA) a prvotného enzýmu kopírujúceho molekulu RNA (RNA polymeráza) v neprítomnosti, ako aj v prítomnosti bunkovej membrány. V neohraničenom priestore je pravdepodobnosť interakcie bielkoviny a RNA veľmi nízka.
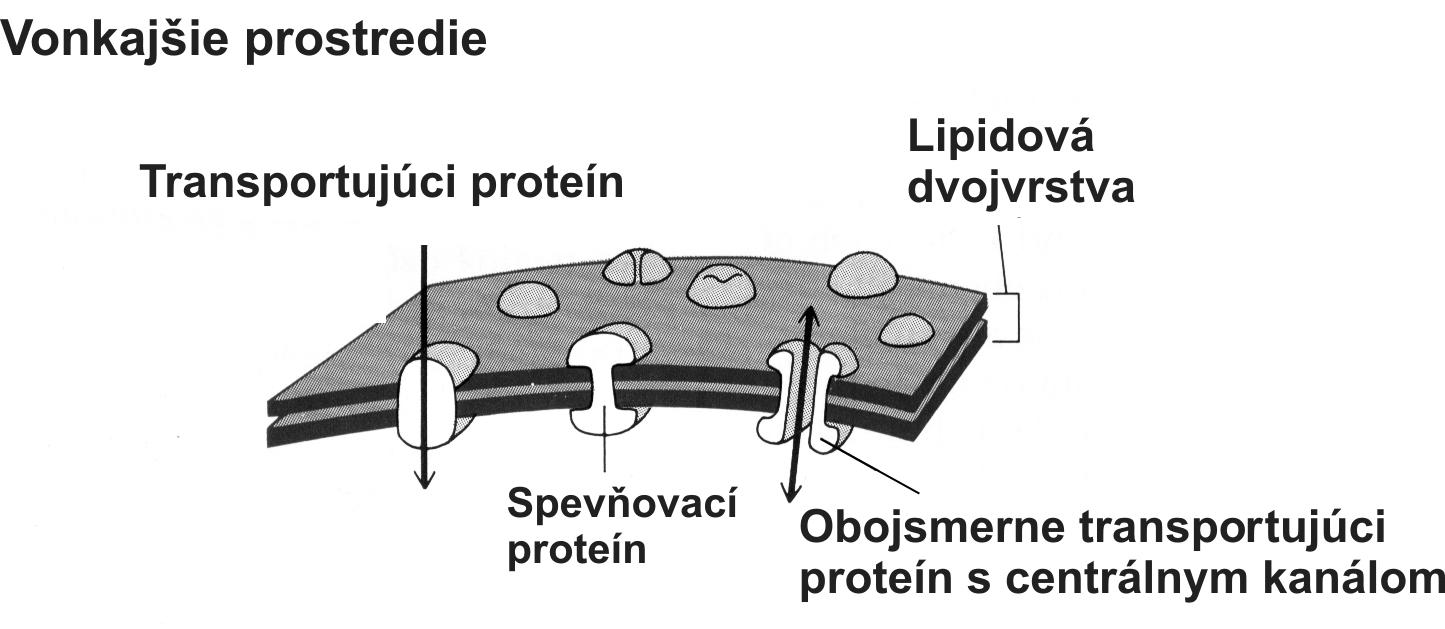
Obr. 7 Po ďalšom zdokonalení bunkovej membrány sa v nej objavujú bielkoviny s transportnými funkciami (prenos solí a živných látok). Transportujúce bielkoviny pracujú buď jednosmerne alebo aj obojsmerne (porov. funkciu ATP syntázy, resp. ATPázy a jeho kanál; opísané v texte).
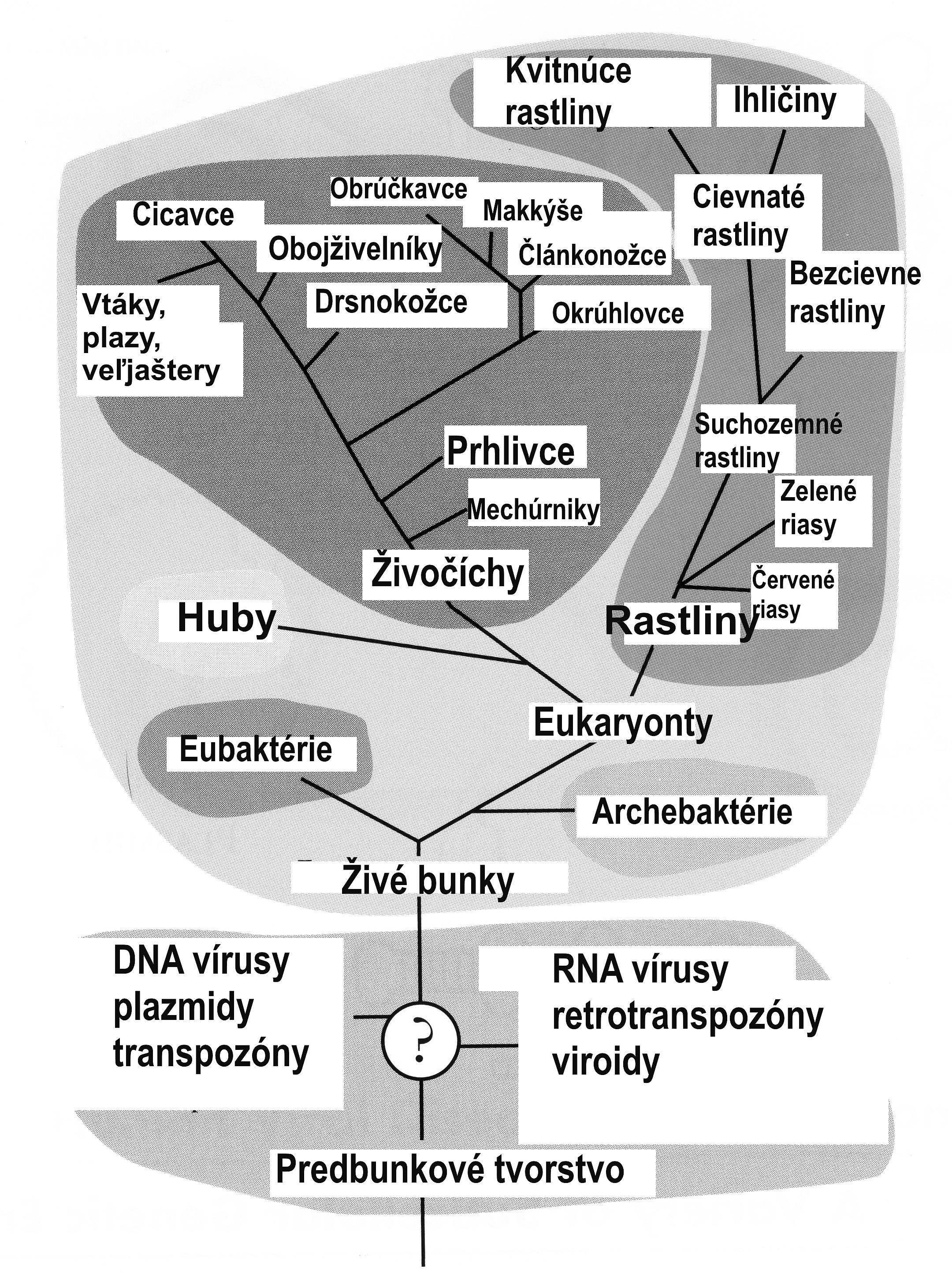
Obr. 8 Strom života. Mnohorakosť tvarov je založená na jednotnom princípe, ktorou je bunková skladba živej hmoty. Jednotný plán s postupným pribúdaním zložitosti umožňuje klasifikáciu organizmov. Eubaktérie sú jednobunkové organizmy, ktoré nemajú jadro. Eukaryonty sú bunky s jadrom; tie po zoskupení sa do mnohobunkových celkov tvoria početné organizmy s veľkou variabilitou tvarov. Klasifikácia organizmov má logický systém preto, že odráža jednotnú koncepciu (program) diferencovaného rozvoja. Je možné logicky očakávať, že ak by bol vývoj organizmov podliehal len a len náhode, nebol by mohol byť jeho výsledok natoľko systematický, aby mohol byť znázornený na schéme. Ak by boli bývali všetky tvary vytvorené súčasne, schéma by nemala tvar stromu, ale skôr hrebeňa. Pri jednom mieste vetvenia (veľjaštery) sa vývoj akoby ocitol v slepej uličke, z ktorej sa ďalší vývoj musel uberať iným smerom. Riešenie bolo závislé od razantného zásahu, ktorý vyvolal hynutie nevídaných rozmerov (teória katastrofy). Bombardovanie planéty malými telesami (od kozmického prachu až po asteroidy) však nepredstavuje nesystémový zásah, lebo samo osebe sa považuje za najpravdepodobnejšie spôsob tvorby planét. Aj voda dopadla na planétu Zem z prstenca ľadových kryštálov, ako dnes obklopujú planétu Saturn.